엡스타인-바 바이러스
"오늘의AI위키"의 AI를 통해 더욱 풍부하고 폭넓은 지식 경험을 누리세요.
1. 개요
엡스타인-바 바이러스(EBV)는 마이클 A. 엡스타인과 이본 바의 이름을 따서 명명되었으며, 전염성 단핵구증, 버킷 림프종, 호지킨 림프종, 위암, 비인두암, 다발성 경화증 등 다양한 질병과 관련된 이중 가닥 DNA 바이러스이다. 주로 B세포와 상피세포에 감염되며, 용해성 복제와 잠복 감염의 두 가지 형태로 존재한다. EBV는 EBV 1형과 EBV 2형으로 나뉘며, EBNA-3 유전자의 차이로 인해 형질전환 능력과 재활성화 능력에서 차이를 보인다. EBV 감염은 ELISA 검사로 진단하며, 아직 백신은 개발되지 않았고, 항바이러스제의 효과는 제한적이다.
더 읽어볼만한 페이지
- 감염성 암원 - 결핵
결핵은 결핵균에 의해 발생하는 감염성 질환으로, 주로 폐에 영향을 미치며 기침, 객혈 등의 증상을 보이지만 무증상일 수도 있고, 호흡기 비말 전파를 통해 감염되며, 항생제 치료를 하지만 약물 내성 문제로 인해 공중 보건 문제로 남아있다. - 감염성 암원 - 자궁경부암
자궁경부암은 인유두종바이러스 감염이 주원인인 자궁 경부의 악성 종양으로, 초기에는 증상이 미미하지만 진행 시 질 분비물 증가, 성교통, 골반통이 나타날 수 있으며, 자궁경부 세포진 검사, 조직 생검 등으로 진단하고 수술, 방사선 치료, 항암 치료 등으로 치료하며, HPV 백신 접종과 정기 검진으로 예방이 가능하다. - IARC 1군 발암 물질 - 토륨
토륨은 은백색의 방사성 악티늄족 금속 원소로, 높은 녹는점과 끓는점을 가지며 지구 지각에 풍부하게 존재하고 핵연료로서의 잠재력을 지니지만, 방사능으로 인해 사용이 감소하고 있다. - IARC 1군 발암 물질 - 황화 카드뮴
황화 카드뮴은 카드뮴 염을 황 이온으로 침전시켜 제조하며, 반도체, 안료 등으로 사용되지만 독성이 있어 흡입 시 위험하고 발암 물질로 분류된다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 라우토카
라우토카는 피지 비치레부섬 서부에 위치한 피지에서 두 번째로 큰 도시이자 서부 지방의 행정 중심지로, 사탕수수 산업이 발달하여 "설탕 도시"로 알려져 있으며, 인도에서 온 계약 노동자들의 거주와 미 해군 기지 건설의 역사를 가지고 있고, 피지 산업 생산의 상당 부분을 담당하는 주요 기관들이 위치해 있다. - 표시 이름과 문서 제목이 같은 위키공용분류 - 코코넛
코코넛은 코코넛 야자나무의 열매로 식용 및 유지로 사용되며, 조리되지 않은 과육은 100g당 354kcal의 열량을 내는 다양한 영양 성분으로 구성되어 있고, 코코넛 파우더의 식이섬유는 대부분 불용성 식이섬유인 셀룰로오스이며, 태국 일부 지역에서는 코코넛 수확에 훈련된 원숭이를 이용하는 동물 학대 문제가 있다.

2. 역사
1964년 마이클 엡스타인과 이본 바가 버킷 림프종 세포에서 엡스타인-바 바이러스를 처음 발견했다.[71][72][102][103] 1961년, 병리학자이자 전자 현미경 전문가였던 엡스타인은 데니스 버킷의 강연을 들었는데, 버킷은 "열대 아프리카에서 가장 흔한 어린이 암—지금까지 인식되지 못한 증후군"에 대해 설명했다. 버킷은 이 강연에서 버킷 림프종의 "유행성 변종" (소아 형태)을 설명했다.[72] 1963년, 우간다에서 배양할 표본이 영국 런던의 미들섹스 병원으로 보내졌다. 배양된 세포에서 바이러스 입자가 확인되었고, 그 결과는 1964년 엡스타인, 버트 아총, 바에 의해 ''더 랜싯''에 발표되었다.[72][73]
엡스타인-바 바이러스(EBV)는 헤르페스바이러스과에 속하는 DNA 바이러스로, B세포와 상피 세포를 포함한 다양한 세포에 감염될 수 있다.[25]
필라델피아 어린이 병원의 베르너 헨레와 게르트루데 헨레는 세포주를 받아 혈청학적 지표를 개발했다.[77] 1967년, 헨레 부부의 실험실 기술자가 단핵구증을 앓았고, 보관된 혈청 샘플을 비교하여 바이러스에 대한 항체가 생성되었음을 확인했다.[73][74][75][104][105][106] 1968년에는 EBV가 감염 후 B 세포를 직접 불멸화시킬 수 있음을 발견하여,[76][107] EBV 관련 감염의 일부 형태를 모방하고,[77] 바이러스와 감염성 단핵구증 간의 연관성을 확인했다.[78][108]
3. 바이러스학
EBV는 지름이 약 122~180nm이며, 약 17만 2천 염기쌍과 85개의 유전자를 가진다.[17][86] 바이러스 DNA는 뉴클레오캡시드라는 단백질 껍질로 둘러싸여 있고, 이는 다시 테구멘트(tegument)라는 단백질 층으로 둘러싸여 있다. 테구멘트는 지질과 당단백질이 포함된 바이러스 외피로 덮여 있다.[22][195]
EBV는 세포 내에서 용해성 복제와 잠복 감염의 두 가지 형태를 가진다. 용해성 복제는 감염성 바이러스 입자(비리온)를 생성하며, B세포에서는 주로 잠복 감염에서 재활성화된 후 일어나지만, 상피 세포에서는 바이러스 침입 후 직접 발생하기도 한다.[22] 잠복 감염은 바이러스 유전자가 숙주 세포 핵 내에 에피솜 형태로 존재하며 제한된 수의 바이러스 유전자만 발현되는 상태이다.[22]
EBV는 B 세포와 상피세포에 모두 감염될 수 있는데, 침투 방식은 서로 다르다. B 세포 침투에는 바이러스 당단백질 gp350과 gp42가, 상피 세포 침투에는 BMRF-2, gH/gL 단백질이 관여한다.[22][27]
EBV는 EBNA-3 유전자의 차이에 따라 1형과 2형으로 나눌 수 있다. 1형은 전 세계 대부분 지역에서 우세하지만, 두 유형 모두 아프리카에서는 흔하게 발견된다.[22]
3. 1. 구조와 유전체
엡스타인-바 바이러스(EBV)는 지름이 약 122~180nm인 DNA 바이러스이며, 약 17만 2천 염기쌍과 85개의 유전자를 가진다.[17][86] 바이러스 DNA는 뉴클레오캡시드라는 단백질 껍질로 둘러싸여 있다. 뉴클레오캡시드는 다시 테구멘트(tegument)라는 단백질 층으로 둘러싸여 있으며, 테구멘트는 지질과 당단백질이 포함된 바이러스 외피로 덮여 있다. 이 외피의 당단백질은 바이러스가 숙주 세포에 침투하는 데 중요한 역할을 한다.[22][195]
3. 2. 친화성
엡스타인-바 바이러스(EBV)는 주로 B세포와 상피 세포에 감염되지만, T세포, NK세포 등 다른 세포에도 감염될 수 있다.[196][85]
EBV의 B세포 감염에는 바이러스 막 당단백질인 gH, gL, gp42로 구성된 복합체가 관여한다. 이 복합체는 B세포 막과의 융합을 매개한다. 그러나 B세포 내에서 조립된 바이러스는 gH/gL/gp42 복합체를 비교적 적게 가지고 있는데, 이는 B세포의 소포체에 있는 사람 백혈구 항원(HLA) MHC 2군에 의해 분해되기 때문이다.[234][26][197]
상피 세포 감염에는 gH/gL 이중 단백질 복합체가 관여하며, 상피 세포 막과의 융합을 매개한다. 상피 세포에서 증식한 바이러스는 gH/gL/gp42 복합체를 모두 갖추고 있다. 상피세포는 HLA 클래스 II 분자를 포함하지 않기 때문에 3중 복합체가 풍부하다.
결과적으로 B세포에서 만들어진 바이러스는 상피 세포에, 상피 세포에서 만들어진 바이러스는 B세포에 더 잘 감염되는 현상이 나타난다. gp42 단백질이 없는 바이러스는 B세포에 결합할 수는 있지만 감염시키지는 못한다.[234][26][197]
3. 3. 복제 주기
EBV는 세포 내에서 용해성 복제와 잠복 감염이라는 두 가지 형태를 가진다.
용해성 복제는 감염성 바이러스 입자(비리온)를 생성하는 과정이다. B세포에서는 주로 잠복 감염에서 재활성화된 후에 일어나지만, 상피 세포에서는 바이러스 침입 후 직접 발생하기도 한다.[22] 용해성 복제 과정에서 바이러스 게놈은 선형화되며, 바이러스 DNA 중합효소가 복제를 담당한다. 용해성 유전자 산물은 즉시-초기, 초기, 후기의 세 단계로 생성된다.[22] 각 단계별 주요 유전자와 기능은 다음 표와 같다.
잠복 감염은 바이러스 유전자가 숙주 세포 핵 내에 에피솜 형태로 존재하며, 제한된 수의 바이러스 유전자만 발현되는 상태이다.[22] 잠복 감염은 발현되는 유전자 패턴에 따라 I형, II형, III형으로 구분된다. B세포에서는 세 가지 형태가 모두 가능하지만, 상피 세포에서는 II형만 가능하다.[33][34] 각 잠복기별 발현 유전자와 생성물은 다음 표와 같다.
B 세포 내 잠복 EBV는 용해성 복제로 전환하기 위해 재활성화될 수 있다. ''생체 내'' 재활성화 유발 요인은 정확히 알려져 있지 않지만, ''시험관 내''에서는 B 세포 수용체 자극으로 재활성화될 수 있다. 따라서 잠복 감염된 B 세포가 관련 없는 감염에 반응한 후 ''생체 내'' 재활성화가 일어날 가능성이 높다.[22]
3. 3. 1. 세포 침투
EBV는 B 세포와 상피세포에 모두 감염될 수 있는데, 이 두 세포에 침투하는 방식은 서로 다르다.B 세포에 침투하기 위해 바이러스 당단백질 gp350은 세포 수용체 CD21(CR2라고도 함)에 결합한다.[27] 그런 다음 바이러스 당단백질 gp42는 세포 MHC II형 분자와 상호 작용한다. 이는 바이러스 외피와 세포막의 지질 이중층 융합을 유발하여 EBV가 B 세포에 진입할 수 있게 한다.[22] 보체 수용체 1(CR1)이라고도 하는 인간 CD35는 gp350/220에 대한 추가적인 부착 인자이며, 미성숙 B 세포를 포함하여 CD21 음성 세포로 EBV가 진입할 수 있는 경로를 제공할 수 있다. EBV 감염은 CD35의 발현을 하향 조절한다.[28]
상피 세포에 침투하기 위해 바이러스 단백질 BMRF-2는 세포 β1 인테그린과 상호 작용한다. 그런 다음 바이러스 단백질 gH/gL은 세포 αvβ6/αvβ8 인테그린과 상호 작용한다. 이는 바이러스 외피와 상피 세포막의 지질 이중층 융합을 유발하여 EBV가 상피 세포에 진입할 수 있게 한다.[22] B 세포 진입과 달리 상피 세포 진입은 바이러스 당단백질 gp42에 의해 실제로 방해를 받는다.[27]
3. 3. 2. 용해성 복제
용균성 주기 또는 생산성 감염은 감염성 비리온의 생성을 초래한다. 엡스타인-바 바이러스(EBV)는 B세포와 상피세포 모두에서 용균성 복제를 겪을 수 있다. B세포에서 용균성 복제는 일반적으로 잠복기로부터 재활성화된 후에만 일어난다. 상피세포에서는 용균성 복제가 종종 바이러스 진입 후 직접적으로 발생한다.[22]용균성 복제가 일어나려면 바이러스 게놈이 선형이어야 한다. 잠복 EBV 게놈은 원형이므로 용균성 재활성화 과정에서 선형화되어야 한다. 용균성 복제 동안 바이러스 DNA 중합효소가 바이러스 게놈 복제를 담당한다. 이는 숙주 세포의 DNA 중합효소가 바이러스 게놈을 복제하는 잠복기와는 대조적이다.[22]
용균성 유전자 산물은 즉시-초기, 초기 및 후기의 세 가지 연속적인 단계로 생성된다.[22]
3. 3. 3. 잠복 감염
용균성 복제와 달리, 잠복기는 바이러스 입자(비리온) 생성을 초래하지 않는다.[22] 대신, 엡스타인-바 바이러스(EBV) 게놈은 원형 DNA 형태로 세포 핵에 에피솜으로 존재하며, 숙주 세포의 DNA 중합 효소에 의해 복제된다.[22] 이는 개인의 기억 B 세포에 지속적으로 존재한다.[17][81] DNA 메틸화 및 세포 크로마틴 구성 요소와 같은 후생 유전적 변화는 잠복 감염된 세포에서 바이러스 유전자의 대부분을 억제한다.[31] EBV 유전자 중 일부만 유전자 발현되어 바이러스의 잠복 상태를 지원한다.[31][17][32]잠복 EBV는 잠복 프로그램으로 알려진 세 가지 패턴 중 하나로 유전자를 발현한다. EBV는 B 세포와 상피 세포 내에서 잠복할 수 있지만, 두 종류의 세포에서 서로 다른 잠복 프로그램이 가능하다.[33][34]
EBV는 세 가지 잠복 프로그램(잠복기 I, 잠복기 II, 잠복기 III) 중 하나를 나타낼 수 있다. 각 잠복 프로그램은 제한적이고 고유한 바이러스 단백질과 바이러스 RNA 세트의 생성을 유도한다.[35][36]
모든 바이러스 단백질 발현이 중단되는 프로그램(잠복기 0)도 존재한다.[37]
B 세포 내에서 세 가지 잠복 프로그램 모두 가능하다.[17] B 세포 내 EBV 잠복기는 일반적으로 잠복기 III, II, I 순으로 진행된다. 각 잠복 단계는 B 세포의 행동에 고유하게 영향을 미친다.[17] 휴지 상태의 미분화 B 세포를 감염시키면, EBV는 잠복기 III로 들어간다. 잠복기 III에서 생성된 단백질과 RNA는 B 세포를 증식하는 블라스트(B 세포 활성화)로 변환시킨다.[17][22] 이후 바이러스는 유전자 발현을 제한하고 잠복기 II로 들어간다. 잠복기 II에서 생성된 더 제한적인 단백질 및 RNA 세트는 B 세포가 기억 B 세포로 분화하도록 유도한다.[17][22] 마지막으로, EBV는 유전자 발현을 더욱 제한하고 잠복기 I로 들어간다. EBNA-1의 발현은 기억 B 세포가 분열할 때 EBV 게놈이 복제되도록 한다.[17][22]
상피 세포 내에서는 잠복기 II만 가능하다.[38]
1차 감염에서 EBV는 구강 인두 상피 세포에서 복제되고 B 림프구에서 잠복기 III, II, I 감염을 확립한다. B 림프구의 EBV 잠복 감염은 바이러스 지속, 상피 세포에서의 후속 복제, 그리고 침으로의 감염성 바이러스 방출에 필요하다. B 림프구의 EBV 잠복기 III 및 II 감염, 구강 상피 세포의 잠복기 II 감염 및 NK- 또는 T-세포의 잠복기 II 감염은 균일한 EBV 게놈 존재 및 유전자 발현이 특징인 악성 종양을 유발할 수 있다.[39]
3. 3. 4. 재활성화
B 세포 내의 잠복 엡스타인-바 바이러스(EBV)는 용해성 복제로 전환하기 위해 재활성화될 수 있다. 이는 ''생체 내''에서 발생하는 것으로 알려져 있지만, 무엇이 이를 유발하는지는 정확히 알려져 있지 않다. ''시험관 내''에서 B 세포 내의 잠복 EBV는 B 세포 수용체를 자극하여 재활성화될 수 있으므로, 잠복 감염된 B 세포가 관련 없는 감염에 반응한 후 ''생체 내'' 재활성화가 일어날 가능성이 높다.[22]특히 용해 감염의 전초기 유전자 BZLF-1은 b-Zip형 전사 인자를 코딩하며, 잠복 상태로부터의 재활성화에 필요하고 충분한 재활성화 책임 유전자이다. 그러나 BZLF-1의 발현은 전사 수준에서 엄격하게 제어되기 때문에 BZLF-1의 전사를 연구하는 것은 EBV 재활성화의 메커니즘을 밝히는 것과 같다. 따라서 전 세계 많은 연구자들이 이 BZLF-1의 프로모터 영역에 주목하여 연구를 진행하고 있다.[203]
실험 환경(in vitro)에서는 잠복 감염 상태에 있는 EBV를 B세포 수용체(면역 글로불린; Ig)를 항Ig 항체로 교차 결합시키는 것 등으로 유도할 수 있으며, 따라서 생체 내 EBV의 재활성화는 잠복 감염 상태에 있는 B세포에 특이적인 항원 자극에 의해 일어날 것으로 생각된다.[85][195] 또한 실험 환경(in vitro)에서는 EBV의 잠복 감염 상태에 있는 B세포를 종양 프로모터 12-O-테트라데카노일포르볼 13-아세테이트·낙산 나트륨에 노출시킴으로써 EBV의 재활성화가 이루어지는 것이 확인되었다.[85]
3. 4. 형질 전환
EBV(엡스타인-바 바이러스)는 B 림프구를 감염시켜 불멸화된, 무한정 분열하는 림프모구 세포주를 형성할 수 있다.[76][40] 이러한 형질 전환에는 EBNA-2, EBNA-3C, LMP-1 단백질이 필수적인 역할을 하는 반면, EBNA-LP와 EBER는 그렇지 않다.[41][204]3. 5. 잠복 항원
EBV 잠복 감염은 잠복 감염 유전자의 발현 패턴에 따라 I형, II형, III형의 3가지로 분류되며, 제한된 종류의 서로 다른 바이러스 단백질, 바이러스 RNA가 생성된다.[200][201]
EBV는 B세포와 상피계 세포에서 잠복 감염될 수 있지만, 잠복 감염 유전자의 발현 패턴은 감염된 세포 종류(B세포 또는 상피계 세포)에 따라 다르다. 또한, 기억 B세포에서의 EBV 감염 양식으로, EBER 이외의 바이러스 유전자 발현이 거의 확인되지 않는 0형이라는 잠복 양식도 존재한다.
B세포에서는 I형, II형, III형 모든 잠복 감염 유전자 발현 패턴이 가능하다.[86] EBV 잠복 감염은 일반적으로 III형, II형, I형 순으로 진행된다. 각 발현 패턴은 B세포의 행동에 특이적인 영향을 미친다.[86] 휴면 중인 나이브 B세포에 감염될 때, EBV는 III형 잠복 감염부터 시작한다. III형 잠복 감염에서 생성되는 단백질과 RNA에 의해 휴면 중인 나이브 B세포는 형질 전환되어 증식성 아구(또는 B세포 활성화)가 된다.[86][195] 그 후, EBV는 잠복 감염 유전자의 발현을 제한하고, II형 잠복 감염으로 돌입한다. II형 잠복 감염에서 발현된 단백질과 RNA는 B세포를 기억 B세포로 분화시킨다.[86][195] 최종적으로 EBV는 잠복 감염 유전자 발현을 더 제한하고, I형 잠복 감염으로 이행한다. I형 잠복 감염에서 생성되는 EBNA-1은 EBV 게놈을 숙주 염색체에 고정하는 앵커로 작용하여, 기억 B세포가 분열할 때 복제될 수 있도록 한다.[86][195]
상피계 세포에서는 II형 잠복 감염만이 가능하다.
I형 잠복 감염은 버킷 림프종, 위암 등에서 보이는 양식으로, EBNA-1과 EBER을 발현한다. II형 감염은 호지킨 림프종, NK/T 림프종, 비인두암, 유방암 등에서 보이며, I형에서 발현하는 유전자에 더하여 LMP-1, LMP-2A, LMP-2B를 발현한다. III형은 기회 감염성 림프종이나, 배양 세포 수준에서 EBV를 B 림프구에 감염시켜 불멸화시킨 경우(림프아구양 세포, LCLs)에 보이며, II형에 더하여 EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-LP 등을 발현한다.
LMP-1은 EBV가 코딩하는 가장 주요한 암 유전자이다. 세포막에 존재하며, CD40의 신호를 모방하여 항시적으로 NF-kB, MAPK, STAT, Akt 등을 활성화함으로써 B세포 증식을 촉진한다. LMP-2A는 B세포 수용체를 모방하여 AKT와 칼슘 신호를 활성화한다. EBER는 단백질을 코딩하지 않은 저분자량 RNA로, RNApol III에 의해 매우 다량으로 전사되므로, ''in situ'' 하이브리다이제이션 등에 의한 바이러스 검출의 마커로 자주 이용된다. EBNA-2는 III형 잠복 감염에서 LMP-1 등의 전사를 증강시킴으로써 불멸화에 관여한다. EBNA-2 자신은 DNA에 결합할 수 없으므로 전사 인자로는 작용할 수 없지만, RBP-Jκ나 PU.1 등 숙주의 전사 인자와 결합함으로써 전사 보조 인자로서 기능한다.
모든 EBV 핵 항원(EBNA)은 EBV 게놈 좌측 단부의 C/W 프로모터 영역에서 시작하는 유전자를 전사한 전령 RNA의 선택적 스플라이싱에 의해 생성된다. 6종류의 EBNA는 EBNA-LP, EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-1 순서로 나열되어 있다.[85] EBNA-1 전사에 특유한 프로모터인 Q 프로모터의 존재도 알려져 있으며, EBV 관련 암세포에서는 주로 이 프로모터가 사용된다. 따라서 대부분의 EBV 관련 암세포에서는 6종류의 EBNA 중 EBNA-1만 발현하고 있다.[85]
EBNA-LP 코딩 영역의 개시 코돈은 해당 유전자를 전사한 전령 RNA의 선택적 스플라이싱에 의해 구성된다. 이 개시 코돈이 없는 상태에서는 EBNA-2, EBNA-3A, EBNA-3B, EBNA-3C, EBNA-1의 각 발현은 이들 유전자 중 어느 것이 선택적으로 스플라이싱되어 해당 EBV 핵 항원으로서 전사되었는지에 따라 결정된다.
3. 6. 유전자/단백질
EBV는 약 172,000 염기쌍을 포함하여 85개의 유전자를 암호화하는 데옥시리보핵산(DNA)의 핵산 이중 나선으로 구성되어 있다.[17] DNA는 단백질 누클레오캡시드로 둘러싸여 있으며, 이는 다시 단백질로 구성된 테구멘트로 둘러싸여 있고, 마지막으로 지질과 당단백질의 표면 돌출부를 모두 포함하는 바이러스 외피로 둘러싸여 있다.[22]바이러스 트로피즘은 EBV가 어떤 세포 유형을 감염시키는지 나타낸다. EBV는 B 세포 및 상피 세포를 포함한 다양한 세포 유형을 감염시킬 수 있다.[25] gHgL gp42의 바이러스 3중 단백질 복합체는 B 세포 막 융합을 매개하며, gHgL의 2중 단백질 복합체는 상피 세포 막 융합을 매개한다. B 세포에서 생성된 EBV는 gHgLgp42 복합체의 수가 적다. 이는 이러한 3중 복합체가 인간 백혈구 항원 MHC 클래스 II와 상호 작용하여 소포체에서 B 세포에 존재하고 분해되기 때문이다. 반대로 상피 세포에서 생성된 EBV는 일반적으로 HLA 클래스 II 분자를 포함하지 않기 때문에 3중 복합체가 풍부하다. 결과적으로, B 세포에서 만들어진 EBV는 상피 세포에 더 잘 감염되며, 상피 세포에서 만들어진 EBV는 B 세포에 더 잘 감염된다. gp42 부분이 없는 바이러스는 인간 B 세포에 결합할 수 있지만 감염시킬 수는 없다.[26]
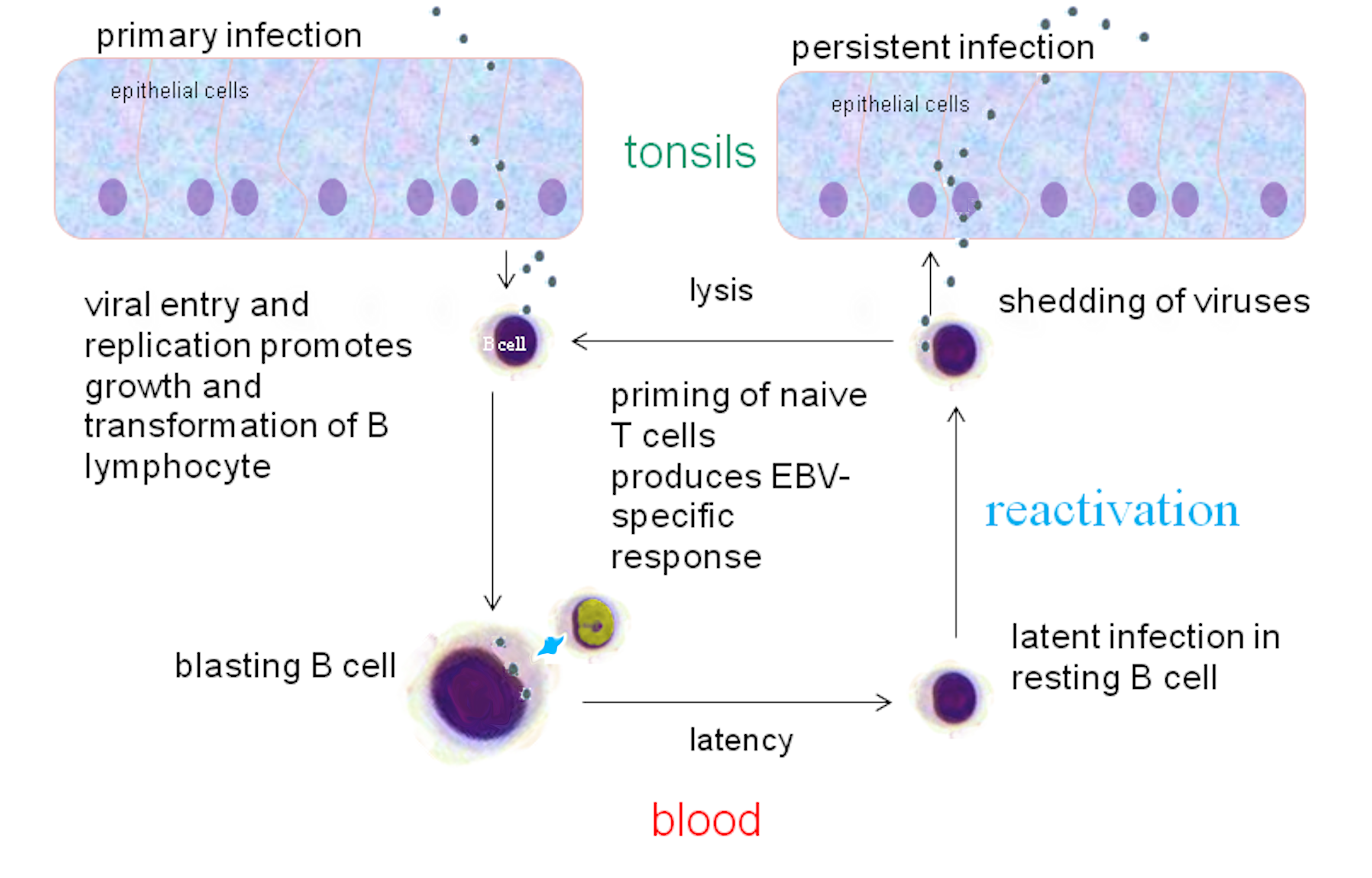
EBV는 B 세포와 상피 세포를 모두 감염시킬 수 있는데, 이 두 세포에 진입하는 메커니즘은 서로 다르다. B 세포에 진입하기 위해 바이러스 당단백질 gp350은 세포 수용체 CD21(CR2)에 결합한다.[27] 그런 다음 바이러스 당단백질 gp42는 세포 MHC II형 분자와 상호 작용한다. 이는 바이러스 외피와 세포막의 지질 이중층 융합을 유발하여 EBV가 B 세포에 진입할 수 있게 한다.[22] 보체 수용체 1(CR1)이라고도 하는 인간 CD35는 gp350/220에 대한 추가적인 부착 인자이며, 미성숙 B 세포를 포함하여 CD21 음성 세포로 EBV가 진입할 수 있는 경로를 제공할 수 있다. EBV 감염은 CD35의 발현을 하향 조절한다.[28]
상피 세포에 진입하기 위해 바이러스 단백질 BMRF-2는 세포 β1 인테그린과 상호 작용한다. 그런 다음 바이러스 단백질 gH/gL은 세포 αvβ6/αvβ8 인테그린과 상호 작용한다. 이는 바이러스 외피와 상피 세포막의 지질 이중층 융합을 유발하여 EBV가 상피 세포에 진입할 수 있게 한다.[22] B 세포 진입과 달리 상피 세포 진입은 바이러스 당단백질 gp42에 의해 실제로 방해를 받는다.[27] EBV가 세포에 진입하면 바이러스 캡시드가 용해되고 바이러스 게놈이 세포 핵으로 수송된다.[29]
용균성 주기 또는 생산성 감염은 감염성 비리온의 생성을 초래한다. 엡스타인-바 바이러스(EBV)는 B 세포와 상피 세포 모두에서 용균성 복제를 겪을 수 있다. B 세포에서 용균성 복제는 일반적으로 잠복기로부터 재활성화된 후에만 일어난다. 상피 세포에서는 용균성 복제가 종종 바이러스 진입 후 직접적으로 발생한다.[22] 용균성 복제가 일어나려면 바이러스 게놈이 선형이어야 한다. 잠복 EBV 게놈은 원형이므로 용균성 재활성화 과정에서 선형화되어야 한다. 용균성 복제 동안 바이러스 DNA 중합효소가 바이러스 게놈 복제를 담당한다. 이는 숙주 세포의 DNA 중합효소가 바이러스 게놈을 복제하는 잠복기와는 대조적이다.[22]
용균성 유전자 산물은 즉시-초기, 초기 및 후기의 세 가지 연속적인 단계로 생성된다.[22] 즉시-초기 용균성 유전자 산물은 전사 활성 인자로 작용하여 후기 용균성 유전자의 유전자 발현을 향상시킨다. 즉시-초기 용균성 유전자 산물에는 BZLF1(Zta, EB1) 및 BRLF1(Rta)이 포함된다.[22] 초기 용균성 유전자 산물은 복제, 대사 및 항원 처리 차단 등 훨씬 더 많은 기능을 한다. 초기 용균성 유전자 산물에는 BNLF2가 포함된다.[22] 마지막으로, 후기 용균성 유전자 산물은 바이러스 캡시드를 형성하는 엡스타인-바 바이러스 바이러스 캡시드 항원(VCA)과 같이 구조적 역할을 하는 단백질인 경향이 있다. BCRF1과 같은 다른 후기 용균성 유전자 산물은 EBV가 면역 체계를 회피하는 데 도움이 된다.[22]
녹차의 폴리페놀인 EGCG는 한 연구에서 시간 및 용량 의존적 방식으로 EBV의 자발적인 용균성 감염을 DNA, 유전자 전사 및 단백질 수준에서 억제하는 것으로 나타났다. EBV 용균성 유전자 ''Zta'', ''Rta'' 및 초기 항원 복합체 EA-D(''Rta''에 의해 유도됨)의 발현은 모든 EBV 감염 단계에서 발견되는 매우 안정적인 ''EBNA-1'' 유전자에는 영향을 미치지 않았다.[30]
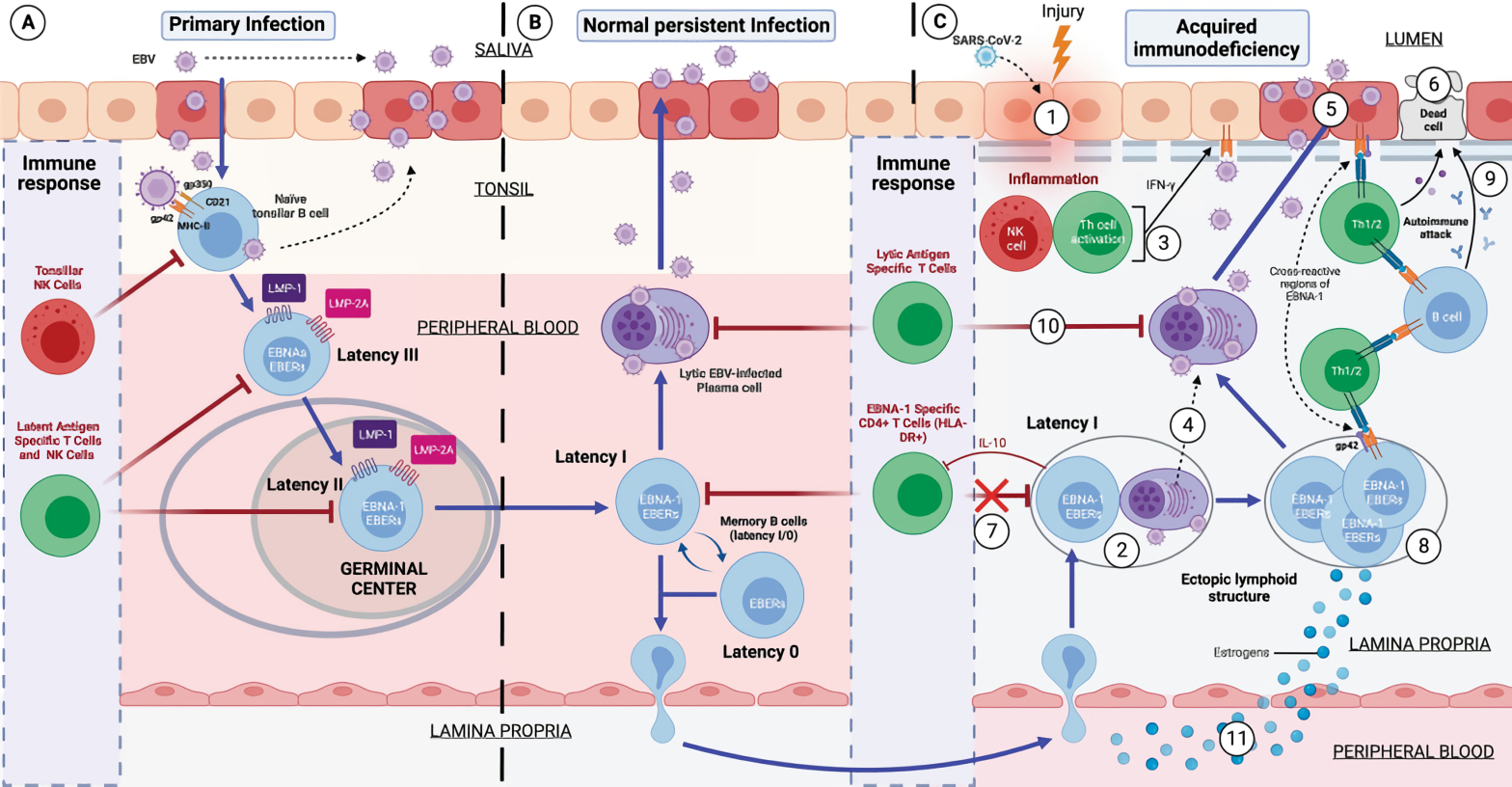
용균성 복제와 달리, 잠복기는 바이리온의 생성을 초래하지 않는다.[22] 대신, EBV 게놈 원형 DNA는 세포 핵에 에피솜으로 존재하며 숙주 세포의 DNA 중합 효소에 의해 복제된다.[22] 이는 개인의 기억 B 세포에 지속적으로 존재한다.[17][81] DNA 메틸화 및 세포 크로마틴 구성 요소와 같은 후생 유전적 변화는 잠복 감염된 세포에서 바이러스 유전자의 대부분을 억제한다.[31] EBV 유전자 중 일부만 유전자 발현되어 바이러스의 잠복 상태를 지원한다.[31][17][32] 잠복 EBV는 잠복 프로그램으로 알려진 세 가지 패턴 중 하나로 유전자를 발현한다. EBV는 B 세포와 상피 세포 내에서 잠복할 수 있지만, 두 종류의 세포에서 서로 다른 잠복 프로그램이 가능하다.[33][34]
EBV는 세 가지 잠복 프로그램(잠복기 I, 잠복기 II, 잠복기 III) 중 하나를 나타낼 수 있다. 각 잠복 프로그램은 제한적이고 고유한 바이러스 단백질과 바이러스 RNA 세트의 생성을 유도한다.[35][36]
모든 바이러스 단백질 발현이 중단되는 프로그램(잠복기 0)도 존재한다.[37] B 세포 내에서 세 가지 잠복 프로그램 모두 가능하다.[17] B 세포 내의 EBV 잠복기는 일반적으로 잠복기 III에서 잠복기 II, 잠복기 I으로 진행된다. 잠복기의 각 단계는 B 세포의 행동에 고유하게 영향을 미친다.[17] 휴지 상태의 미분화 B 세포를 감염시키면, EBV는 잠복기 III로 들어간다. 잠복기 III에서 생성된 단백질과 RNA 세트는 B 세포를 증식하는 블라스트(B 세포 활성화)로 변환시킨다.[17][22] 나중에 바이러스는 유전자 발현을 제한하고 잠복기 II로 들어간다. 잠복기 II에서 생성된 더 제한적인 단백질 및 RNA 세트는 B 세포가 기억 B 세포로 분화하도록 유도한다.[17][22] 마지막으로, EBV는 유전자 발현을 더욱 제한하고 잠복기 I으로 들어간다. EBNA-1의 발현은 기억 B 세포가 분열할 때 EBV 게놈이 복제되도록 한다.[17][22]
상피 세포 내에서는 잠복기 II만 가능하다.[38] 1차 감염에서 EBV는 구강 인두 상피 세포에서 복제되고 B 림프구에서 잠복기 III, II, I 감염을 확립한다. B 림프구의 EBV 잠복 감염은 바이러스 지속, 상피 세포에서의 후속 복제, 그리고 침으로의 감염성 바이러스 방출에 필요하다. B 림프구의 EBV 잠복기 III 및 II 감염, 구강 상피 세포의 잠복기 II 감염 및 NK- 또는 T-세포의 잠복기 II 감염은 균일한 EBV 게놈 존재 및 유전자 발현이 특징인 악성 종양을 유발할 수 있다.[39] EBV가 B 림프구를 감염시키면 이 세포들이 "불멸화"되어 바이러스가 세포가 무한정 분열하도록 만든다.[76]
EBV가 ''생체 외''에서 B 세포를 감염시키면, 결국 무한정 증식할 수 있는 림프모구 세포주가 나타난다. 이 세포주의 성장 변환은 바이러스 단백질 발현의 결과이다.[40] EBNA-2, EBNA-3C, LMP-1은 변환에 필수적인 반면, EBNA-LP와 EBER는 그렇지 않다.[41] EBV에 의한 자연 감염 이후, 바이러스는 지속적인 감염을 확립하기 위해 유전자 발현 프로그램의 일부 또는 전부를 실행하는 것으로 생각된다. 숙주 면역이 처음에는 부재하므로, 용해성 주기는 숙주 내의 다른 (아마도) B 림프구를 감염시키기 위해 많은 수의 비리온을 생성한다.
잠복 프로그램은 감염된 B 림프구를 재프로그래밍하고 전복하여 증식시키고 감염된 세포를 바이러스가 지속적으로 존재한다고 추정되는 부위로 가져간다. 결국, 숙주 면역이 발달하면 바이러스는 유전자 대부분(또는 아마도 전부)을 끄고 가끔 재활성화되어 자손 비리온을 생성함으로써 지속된다. EBV에 의한 인체의 후생 유전학적 조작은 세포의 게놈을 변경하여 종양 유발 표현형을 남길 수 있다.[82]
EBV의 지속 부위는 골수일 수 있다. 자신의 골수가 EBV 음성 기증자의 골수로 대체된 EBV 양성 환자는 장기 이식 후 EBV 음성으로 밝혀졌다.[43] 모든 EBV 핵 단백질은 게놈의 왼쪽 끝에 있는 Cp 또는 Wp 프로모터에서 시작하는 전사체의 선택적 스플라이싱에 의해 생성된다. 유전자들은 게놈 내에서 ''EBNA-LP/EBNA-2/EBNA-3A/EBNA-3B/EBNA-3C/EBNA-1'' 순서로 배열되어 있다.
EBNA-LP 코딩 영역의 개시 코돈은 핵 단백질 전사체의 대체 스플라이스에 의해 생성된다. 이 개시 코돈이 없으면, 이러한 유전자 중 어떤 것이 전사체로 선택적으로 스플라이싱되는지에 따라 ''EBNA-2/EBNA-3A/EBNA-3B/EBNA-3C/EBNA-1''이 발현된다.
{| class="wikitable"
|-
! 단백질/유전자/항원
! 시기
! 설명
|-
| '''EBNA-1'''
| 잠복기+용해기
| EBNA-1 단백질은 바이러스 게놈 내의 복제 기점(oriP)에 결합하여 숙주 세포 분열 동안 에피솜의 복제 및 분리를 매개한다. 이는 그룹 I 잠복기 동안 발현되는 유일한 바이러스 단백질이다.
|-
| '''EBNA-2'''
| 잠복기+용해기
| EBNA-2는 주요 바이러스 전사 활성 인자이다.
|-
| '''EBNA-3'''
| 잠복기+용해기
| 이 유전자들은 또한 숙주 RBP-Jκ 단백질에 결합한다.
|-
| '''LMP-1'''
| 잠복기
| LMP-1은 EBV 매개 성장 형질 전환에도 필수적인 6개의 스팬을 가진 막횡단 단백질이다.
|-
| '''LMP-2'''
| 잠복기
| LMP-2A/LMP-2B는 티로신 키나아제 신호를 차단하는 작용을 하는 막횡단 단백질이다.
|-
| '''EBER'''
| 잠복기
| EBER-1/EBER-2는 특정 핵단백질 입자에 결합하는 소형 핵 RNA로, PKR(dsRNA-의존적 세린/트레오닌 단백질 키나아제)에 결합하여 기능을 억제할 수 있다. EBER는 EBV 감염 세포에서 전사되는 가장 풍부한 EBV 산물이다. 이들은 조직학적 조직에서 EBV를 검출하는 표
3. 7. EBV의 유형
EBV는 EBV 1형과 EBV 2형, 두 가지 주요 유형으로 나눌 수 있다. 이 두 아형은 서로 다른 EBNA-3 유전자를 가지고 있어 형질전환 능력과 재활성화 능력에서 차이를 보인다. 1형은 전 세계 대부분에서 우세하지만, 두 유형 모두 아프리카에서 동일하게 널리 퍼져 있다. 제한 효소로 바이러스 게놈을 절단하고, 그 결과로 생성된 분해 패턴을 젤 전기영동으로 비교하여 EBV 1형과 EBV 2형을 구별할 수 있다.[22]4. 질병
EBV는 전염성 단핵구증을 유발한다.[50] EBV 감염은 어린이에게는 증상이 거의 없거나 없을 수 있지만, 청소년기나 성인기에 감염되면 피로, 발열, 인두 염증, 목 림프절의 비대, 비장 비대, 간 비대, 발진을 일으킬 수 있다.[18] 만성 피로 증후군도 EBV 감염과 관련이 있다.[51][52]
EBV는 버킷 림프종,[53] 혈구탐식성 림프조직구증,[54] 호지킨 림프종,[55] 위암,[56][57] 비인두암,[58] 다발성 경화증,[14][15][59][60] 림프종양 육아종증 등 여러 질병과 관련이 있다.[61] 2022년 1,000만 명의 군인을 대상으로 한 연구에서는 EBV 감염이 다발성 경화증 위험을 30배 이상 증가시킨다는 사실이 밝혀졌다.[62]
EBV는 지안노티-크로스티 증후군, 다형성 홍반, 급성 생식기 궤양, 구강 모상 백반증과도 관련이 있다.[63] 또한, 심각한 과민성 모기 물림 알레르기 반응,[64] 엡스타인-바 바이러스 양성 점막 피부 궤양, 수두 유사 림프증식성 질환 같은 비악성 림프증식성 질환과 엡스타인-바 바이러스 양성 버킷 림프종,[65] 엡스타인-바 바이러스 양성 호지킨 림프종,[66] 원발 삼출 림프종 같은 악성 림프증식성 질환의 발달과도 관련이 있다.[67]
EBV는 파킨슨병, 루이 소체 치매, 다계통 위축증 등 알파-시누클레인 응집 관련 질환과도 관련이 있다.[68] EBNA1은 11번 염색체 특정 영역에 염색체 파괴를 유도하여 백혈병 관련 유전자 변형을 일으킬 수 있다.[69] EBV의 지속성과 숙주 게놈 통합은 외투 세포 림프종 등 악성 종양에서 발견되며, 이는 EBV가 종양 형성에 영향을 미치는 기전을 보여준다.[70]
4. 1. 감염성 단핵구증
어린이는 엡스타인-바 바이러스(EBV)에 감염되어도 대부분 증상을 나타내지 않지만, 청소년이나 성인이 되어서 처음 감염되면 피로, 발열, 인후염, 목의 임파선염, 비장종대, 간비대, 발진 등의 증상이 나타난다.[231][50][18] 만성 피로 증후군 또한 EBV 감염과 관련이 있다.[51][52]4. 2. 악성 종양
EBV는 종양 바이러스로서 다양한 암 발생에 관여한다.[53][55][56][57][58] EBV 양성 암은 다단계 발암 과정을 거치며, EBV 유전자의 암 유전자 효과, 숙주 게놈의 유전적/후성유전학적 변화, 면역계의 영향 등이 관여한다.EBV는 버킷 림프종,[53] 호지킨 림프종,[55] 비인두암,[58] 위암,[56][57] T/NK 림프종, 평활근육종,[112][113] 타액선암,[116][85] 유방암[88] 등과 관련이 있다.
비인두암은 중국 남부 지역에서 높은 발병률을 보이며, 거의 100%의 종양 세포에서 EBV가 검출된다. 소금에 절인 생선 섭취와도 관련이 있다.[58] 대한민국에서는 위암의 약 10% 미만에서 EBV가 검출된다. 에이즈나 장기 이식 환자에서 EBV 양성 평활근육종 발생 보고가 있다.[114][115][85] 인도,[117] 중국,[118][119] 북아프리카,[120][121] 남유럽[122][123][124] 등에서 EBV와 유방암의 관련성이 보고되었다.
EBV는 EBNA1을 통해 11번 염색체의 특정 영역에 파괴를 유도할 수 있으며, 이는 백혈병과 관련된 유전자의 변형과 관련이 있을 수 있다.[69] EBV의 지속성과 숙주 게놈으로의 통합은 외투 세포 림프종을 포함한 악성 종양에서 관찰되며, 이는 EBV가 종양 형성에 영향을 미치는 메커니즘을 시사한다.[70]
4. 2. 1. B세포 림프종
엡스타인-바 바이러스(EBV)가 B 림프구를 감염시키면, 이 세포들은 불멸화되어 무한정 분열하게 된다. 일반적인 세포는 수명이 제한되어 있지만, EBV에 감염된 B 림프구는 세포의 행동이 바뀌어 평소보다 훨씬 오래 분열하고 생존하여 "불멸"하게 된다. 이를 통해 바이러스는 개인의 평생 동안 신체 내에 지속적으로 존재할 수 있다.[76]''생체 외''에서 EBV에 B 세포가 감염되면, 무한정 증식할 수 있는 림프모구 세포주가 나타난다. 이 세포주의 성장 변환은 바이러스 단백질 발현의 결과이다.[40] EBNA-2, EBNA-3C, LMP-1은 변환에 필수적인 반면, EBNA-LP와 EBER는 그렇지 않다.[41]
EBV 자연 감염 이후, 바이러스는 지속적인 감염을 확립하기 위해 유전자 발현 프로그램의 일부 또는 전부를 실행하는 것으로 추정된다. 초기에는 숙주 면역이 없으므로, 용해성 주기는 많은 수의 비리온을 생성하여 숙주 내의 다른 B 림프구를 감염시킨다.
잠복 프로그램은 감염된 B 림프구를 재프로그래밍하고 전복하여 증식시키고, 감염된 세포를 바이러스가 지속적으로 존재하는 부위로 이동시킨다. 숙주 면역이 발달하면 바이러스는 대부분의 유전자를 끄고 가끔 재활성화되어 자손 비리온을 생성함으로써 지속된다. 바이러스 재활성화와 바이러스 유전자 발현을 활성화하는 세포를 제거하는 숙주 면역 감시 사이의 균형이 이루어진다. EBV에 의한 인체의 후생 유전학적 조작은 세포의 게놈을 변경하여 종양 유발 표현형을 남길 수 있다.[82] 결과적으로 EBV에 의한 변형은 숙주가 EBV 관련 암을 발병할 가능성을 증가시킨다.[83] EBV 관련 암은 후생 유전학적 변화를 자주 일으키지만 돌연변이를 일으킬 가능성은 적다는 점에서 독특하다.[42]
EBV의 지속 부위는 골수일 수 있다. EBV 양성 환자가 자신의 골수를 EBV 음성 기증자의 골수로 대체하는 장기 이식을 받은 후 EBV 음성으로 밝혀진 사례가 있다.[43]
4. 2. 2. 상피세포암
비인두암(nasopharyngeal carcinoma; NPC)은 중국 남부에서 많이 발생하며, 거의 모든 경우(100%) 종양 세포에서 엡스타인-바 바이러스(EBV)가 검출되므로 EBV가 주원인으로 생각된다. 비인두 상부에는 편도가 있어 해부학적 위치상 EBV 양성 림프구와 EBV에 노출될 기회가 높다. EBV 외에도 소금에 절인 생선 섭취와 강한 상관관계가 있으며, 생선에 함유된 니트로소아민 등의 변이원성 물질이 관여하는 것으로 추정된다. 림프구 침윤이 심해 이전에는 림프상피종(lymphoepithelioma)이라고 불렸지만, 암의 본질은 림프구가 아닌 상피 세포이다. Rassif1A, P16, TP53, Cyclin D1 등의 변이가 동반되는 것으로 보고되었다.위암(gastric carcinoma; GC)의 약 10% 미만에서 EBV가 검출된다. EBV 양성 위암은 상인두암과 마찬가지로 거의 모든 경우에 림프구 침윤이 심하게 동반된다. TP53, ARID1의 변이가 보고되었으며, P16이나 E-cadherin 등의 침묵 현상도 EBV 양성 위암과 관련이 있는 것으로 밝혀졌다.
4. 3. 자가면역질환
EBV는 다발성 경화증, 전신 홍반 루푸스, 류마티스 관절염, 쇼그렌 증후군, 강직성 척추염, 전신 경화증, 피부근염, 원발성 담즙성 담관염, 원발성 경화성 담관염, 궤양성 대장염, 크론병, 건선, 백반증, 수포성 유사천포창, 원형 탈모증, 특발성 확장성 심근증, 1형 당뇨병, 바세도우병, 하시모토병, 중증 근무력증, IgA 신병증, 막성 신병증, 악성 빈혈 등 다양한 자가면역질환과 관련이 있다.[90]호주의 퀸즐랜드 대학교 의과대학의 마이클 P. 펜더(Michael P. Pender)는 EBV에 의한 자가면역 질환 발병 메커니즘 가설을 제시했다.[125][126][90] 이 가설은 "펜더의 가설(Pender's hypothesis)"이라고 불리며, EBV 감염, CD8+ 세포독성 T세포 기능 부전, 비타민 D 결핍 등이 자가면역질환 발병에 관여하는 기전을 제시한다.
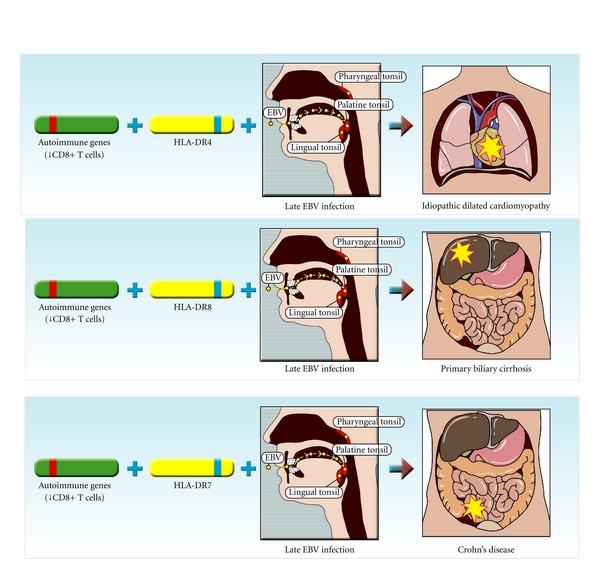
펜더의 가설은 유전 등으로 EBV에 대한 CD8+T세포 반응에 부전이 일어나, EBV 감염 자기 반응성 기억 B세포가 항원 제시 세포로 작용하여 자기 항원 T세포 인식이 가능하게 되어 자가 면역 반응이 생긴다는 것이다.[90]
오사카 대학 미생물 병연구소/면역학 프론티어 연구센터·하버드 대학교 연구 그룹은 2015년, EBV 잠복 감염 유전자 LMP-2A가 B세포 수용체 신호를 모방하고, 형질 세포 분화 촉진 인자(Zbtb20)가 출현하여 자기 반응성 B세포가 생존, 자가면역 질환을 유발한다는 것을 밝혀냈다.[91][92][93]
돗토리 대학 의과대학 분자 병리학 분야 연구 그룹은 2017년, EBV 감염 B세포로부터 바세도우병 자가항체인 항 갑상선 자극 호르몬 수용체 항체(TRAb)가 생산되는 메커니즘을 밝혀냈다. EBV 잠복 감염 유전자 LMP-1이 T세포 비의존적 CD40 신호를 모방, NF-κB 활성화, 활성화 유도 시티딘 탈아미노 효소(AID) 발현 촉진, 클래스 스위칭을 가능하게 하여 다클론성 항체를 생산한다. EBV 감염 B세포가 자기 반응성 항체 가변부를 가지면 자가항체를 생산할 수 있다.[94][95]
류마티스 관절염의 경우, EBNA-1 글리신-알기닌 반복 구조 알기닌이 시트룰린으로 변환, viral citrullinated peptides가 되고, 류마티스 관절염(RA) 특이 자가항체 항 cyclic citrullinated peptide(CCP) 항체와 교차 반응을 일으킨다고 보고되었다.[133]
2012년 쓰루미 대학 치학부 연구 그룹은 다이옥신 일종인 TCDD가 EBV를 활성화, 쇼그렌 증후군 발병에 관여한다고 발표했다.[134]
다발성 경화증의 경우, 2018년 현재 펜더의 가설은 검증 단계에 있지만, 환자에게서 EBV에 대한 T 세포 반응 피로가 일어난다는 것이 밝혀졌다.[140][141]
염증성 장 질환(IBD)의 경우, 최근 일본에서도 EBV와의 관련성이 밝혀지고 있다.
4. 4. 기타 질환
엡스타인-바 바이러스(EBV)는 만성 활동성 EBV 감염증(CAEBV), α-시누클레인 응집 관련 질환, 이상한 나라의 앨리스 증후군, 급성 신부전 등과 같은 여러 질병과 관련이 있다.만성 활동성 EB 바이러스 감염증(CAEBV)은 EBV에 감염된 NK 세포나 T 세포가 증식하는 질환이다. "만성 활동성"이라는 이름은 항 바이러스 캡시드 항원(VCA)-IgG, 항 초기 항원(EA)-IgG와 같이 용해 감염과 관련된 유전자에 대한 항체 수치가 높은 경우가 많기 때문에 붙여졌지만,[179] 증식하는 T/NK 세포에서 EBV는 용해 감염이 아닌 잠복 상태에 있다. 전염성 단핵구증에서는 EBV가 B세포를 감염시키지만, CAEBV에서는 NK 세포나 T세포가 감염된다. CAEBV 환자의 조직에서는 EBER-1의 ''in situ'' 하이브리다이제이션을 통해 많은 EBV 감염 림프구를 확인할 수 있으며, 환자의 말초 혈액 단핵구(PBMC)에서 EBV 게놈이 높게 나타나는 경우가 많다.[179]
전염성 단핵구증과 비슷한 증상이 오랫동안 지속되는 것 외에도, 모기 물린 자국에 과민 반응을 보이거나, 적혈구 탐식 증후군 등이 함께 나타나는 경우가 있다. 일본을 포함한 동아시아에서 비교적 많이 발생한다. 일부 경우에는 만성 활동성 EB 바이러스 감염증이 악성으로 변하여 T/NK 림프종이 발생하기도 한다. EBV에 감염된 주된 세포가 T 세포인지 NK 세포인지에 따라 치료 결과가 달라진다.[179]
EBV 감염 세포가 CD3+ 세포인 경우 T 세포형으로 분류하며, CD4+ 세포와 CD8+ 세포로 나뉜다. EBV에 의한 적혈구 탐식 증후군의 경우, EBV에 감염된 림프구는 대부분 CD8+ 세포이며, CAEBV에서는 주로 CD4+ 세포에 감염되는 경우가 많다. 특히 활성화된 T 세포에서 많은 EBV 감염 세포가 확인된다. CD3-이면서 CD16+ 또는 CD56+ 세포에 EBV가 감염된 경우에는 NK 세포형으로 분류한다.[179]
T 세포형 CAEBV는 고열과 VCA-IgG, EA-IgG 항체 수치가 높은 것이 특징이다. 이는 EBV에 감염된 T 세포가 활성화되어 인터페론 감마, IL-6, TNF-α 등의 염증성 사이토카인을 방출하여 심각한 염증과 발열을 일으키는 것으로 생각된다.[179] 반면 NK 세포형 CAEBV는 HMB, 대과립 림프구 증가증, IgE 항체 수치가 높은 것이 특징이다.[179]
EBV는 B 세포에서 CD40L 발현을 유발하고, CD40과 CD40L의 공동 발현을 일으킨다.[180] 이 두 분자 간의 상호 작용은 공동 자극(co-stimulation)에 의한 세포 생존 신호를 냄으로써 B 세포 형질 전환에 큰 역할을 한다. EBV 감염에 의한 CD40과 CD40L 공동 발현은 B 세포뿐만 아니라 T/NK 세포의 증식에도 영향을 주어, T 세포나 NK 세포의 불멸화를 촉진할 수 있다.[181]
파킨슨병, 레비 소체 치매, 다계통 위축증과 같은 α-시누클레인 응집이 관여하는 신경 퇴행성 질환과 EBV의 관련성이 지적되고 있다. EBV의 잠복 감염 유전자 산물인 LMP-1에 대한 항체가 α-시누클레인에 대해 교차 반응(크로스 리액션)을 일으킨다는 것이 밝혀졌다.[182]
이상한 나라의 앨리스 증후군(AIWS)은 지각된 외부 사물의 크기나 자신의 신체 크기가 평소와 다르게 느껴지는 것을 주요 증상으로 하며, 다양한 주관적인 이미지의 변용을 일으키는 증후군이다. 이 증상은 엡스타인-바 바이러스(EBV)의 첫 감염으로 유발된 중추신경계에서의 보고가 많다.[183][184] EB 바이러스에 감염된 환자에게서 제한적인 영상법으로 단기간 동안 일시적인 대뇌 피질의 광범위한 변이가 관찰되었다는 보고가 있지만,[185] 국소적인 병소를 보이는 보고는 없으며, 뇌의 넓은 범위가 관여하는 것으로 시사된다.
EBV의 첫 감염과 함께 급성 신부전을 일으키는 경우가 있다. 원인을 알 수 없는 급성 신부전을 겪는 소아의 경우, EBV를 원인으로 고려해야 한다.[186]
5. 진단
EBV 감염 진단에는 주로 효소 결합 면역 흡착 분석법(ELISA)을 이용한 혈청학적 검사가 사용된다. 이 검사를 통해 EBV DNA에 의해 암호화된 항체(IgM 및 IgG)를 감지한다.[48] VCA-IgM, VCA-IgG, EA, EBNA 항체 유무를 확인하여 감염 여부 및 시기를 판단할 수 있다.
중합 효소 연쇄 반응(PCR)을 통해 EBV 게놈의 존재를 직접 검출할 수도 있지만, 이 방법은 바이러스에 대한 면역 체계의 반응을 반영하지 않기 때문에 임상적 유용성은 제한적이다. EBV 바이러스 부하는 감염의 임상 증상과 잘 연관되지 않는다.[49]
조직학적 검사에서는 엡스타인-바 바이러스(EBV)로 암호화된 작은 RNA(EBER)가 EBV에 감염된 세포에서 전사되는 가장 풍부한 EBV 산물이므로, EBER in situ hybridization이 EBV 감염 세포 검출에 사용된다.[44]
6. 예방 및 치료
엡스타인-바 바이러스(EBV)는 비교적 복잡한 바이러스이므로 아직 완전히 밝혀지지 않았다. 전 세계 연구소에서 이 바이러스를 지속적으로 연구하고 있으며, 이 바이러스가 유발하는 질병을 치료할 새로운 방법을 개발하고 있다. 엡스타인-바 바이러스를 ''시험관 내''에서 연구하는 한 가지 일반적인 방법은 박테리아 인공 염색체를 사용하는 것이다.[79] 엡스타인-바 바이러스는 지속적인 잠복 상태로 실험실에서 유지하고 조작할 수 있으며, 유전체 연구를 통해 용해성 재활성화 및 잠복 바이러스 에피솜의 조절을 탐구할 수 있었다.[80]
단순 헤르페스 또는 대상 포진 치료에 승인된 항바이러스제인 발라시클로비르를 장기간 사용하면 엡스타인-바 바이러스를 제거할 수 있을지 모른다는 이론이 있지만, 그러한 이론은 어떠한 연구로도 확인되지 않았다.[82] 항바이러스제는 바이러스 DNA 복제를 억제하여 작용하지만, 엡스타인-바 바이러스에 효과가 있다는 증거는 거의 없다. 더욱이, 이러한 약물은 비싸고, 항바이러스제 내성을 유발할 위험이 있으며, 1%에서 10%의 경우 불쾌한 부작용을 유발할 수 있다.[83]
6. 1. 백신
현재까지 EBV에 대한 백신은 개발되지 않았다.[100] 2006년부터 2008년까지 몇 차례의 임상 시험이 진행되었다.[187][188][189] EBV의 gp350 등이 백신의 표적으로 지지받아 왔다.[190][189] 또한, EBV의 바이러스 유사 입자 (VLP)를 이용한 EBV 백신 개발이 활발하게 이루어지고 있다.[191]EBV 백신 개발의 어려움은 EBV가 잠복 감염과 용해 감염에서 완전히 다른 바이러스 단백질을 발현한다는 점에 있다. 용해 감염 유전자의 전사 산물 (gp350 등)을 표적으로 한 백신을 만들더라도, 잠복 감염 시에는 발현되지 않기 때문이다.
특히, EBV가 감염된 B 세포 내에서 잠복 감염의 I형을 띠고 있을 때는 EBNA-1과 EBERs만 발현되기 때문에 CD8+세포 독성 T 세포가 해당 EBV 감염 B 세포를 제거하기 매우 어렵다.
6. 2. 항바이러스제
아시클로비르 등 항헤르페스 바이러스제는 엡스타인-바 바이러스(EBV)에 대한 효과가 제한적이다.[193] 만성 활동성 EBV 감염증 치료의 하나로 아시클로비르를 대량 투여하기도 하지만, 이 역시 효과는 제한적이다.[85] 과거에는 EBV에 효과적인 항바이러스제인 소리부딘이 있었으나[192][193], 항암제 5-FU와의 병용으로 인한 약화 사고 발생으로 현재는 판매가 중단되었다. 소리부딘의 대사산물인 브로모비닐우라실(BVU)은 5-FU의 대사 효소인 DPD(dihydropyrimidine dehydrogenase)와 결합하여 5-FU의 혈중 농도를 높여 부작용을 일으켰다.
소리부딘과 유사한 항바이러스제인 브리부딘 또한 EBV에 효과적이지만[193], 소리부딘 약화 사고 이후 일본에서는 브리부딘의 대사 산물이 소리부딘과 같은 BVU이므로 사용되지 않고 있다.
참조
[1]
웹사이트
ICTV Taxonomy history: 'Human gammaherpesvirus 4'
https://ictv.global/[...]
2019-01-10
[2]
논문
Beyond cytomegalovirus and Epstein–Barr virus: A review of viruses composing the blood virome of solid organ transplant and hematopoietic stem cell transplant recipients
[3]
간행물
Urgency and necessity of Epstein–Barr virus prophylactic vaccines.
2022
[4]
논문
Epstein–Barr virus (EBV)-associated lymphoid proliferations, a 2018 update
2018-09
[5]
논문
Spectrum of Epstein–Barr virus-related diseases: A pictorial review
2009-01
[6]
논문
Oral manifestations in the era of HAART
2003-02
[7]
논문
Alice in Wonderland Syndrome: A clinical and pathophysiological review
[8]
논문
Post-infectious acute cerebellar ataxia in children
2003-09
[9]
논문
Epstein–Barr virus in autoimmune diseases
2008-10
[10]
논문
Autoimmune disease: A role for new anti-viral therapies?
2011-12
[11]
논문
CD8+ T cell deficiency, Epstein–Barr virus infection, vitamin D deficiency, and steps to autoimmunity: A unifying hypothesis
[12]
논문
Global and regional incidence, mortality and disability-adjusted life-years for Epstein–Barr virus-attributable malignancies, 1990–2017
2020-08
[13]
논문
Epstein–Barr virus and multiple sclerosis
2022-01
[14]
논문
Longitudinal analysis reveals high prevalence of Epstein–Barr virus associated with multiple sclerosis
American Association for the Advancement of Science (AAAS)
2022-01
[15]
논문
Epstein-barr virus infection and multiple sclerosis: a review
2010-09
[16]
논문
Epstein–Barr virus is present in the brain of most cases of multiple sclerosis and may engage more than just B cells
[17]
논문
Reactivation of Epstein–Barr virus from latency
2004-11
[18]
웹사이트
About 90% of adults have antibodies that show that they have a current or past EBV infection
https://www.cdc.gov/[...]
US CDC
2020-09-28
[19]
논문
NIH conference. Epstein–Barr virus infections: Biology, pathogenesis, and management
1993-01
[20]
웹사이트
Epstein–Barr virus and infectious Mononucleosis
https://www.cdc.gov/[...]
U.S. Centers for Disease Control and Prevention (CDC)
2011-12-29
[21]
논문
Is EBV persistence in vivo a model for B cell homeostasis?
1996-08
[22]
논문
Progress and problems in understanding and managing primary Epstein–Barr virus infections
2011-01
[23]
간행물
Scientists uncover first atomic structure of Epstein–Barr virus nucleocapsid
https://phys.org/new[...]
2020-10-04
[24]
논문
CryoEM structure of the tegumented capsid of Epstein–Barr virus
2020-10
[25]
논문
Epstein Barr virus entry; kissing and conjugation
2014-02
[26]
논문
Epstein–Barr virus lacking glycoprotein gp42 can bind to B cells but is not able to infect
1998-01
[27]
웹사이트
Entrez gene: CR2 complement component (3d/Epstein Barr virus) receptor 2
https://www.ncbi.nlm[...]
[28]
논문
Human complement receptor type 1 / CD35 is an Epstein–Barr Virus receptor
2013-02
[29]
논문
Epigenetic lifestyle of Epstein–Barr virus
2020-04
[30]
논문
(−)-Epigallocatechin-3-gallate inhibition of Epstein–Barr virus spontaneous lytic infection involves ERK1/2 and PI3-K/Akt signaling in EBV-positive cells
2013-03
[31]
논문
Epigenetic lifestyle of Epstein–Barr virus
2020-04
[32]
논문
Epstein–Barr virus and the B cell: That's all it takes
1996-05
[33]
논문
The effects of the Epstein–Barr virus latent membrane protein 2A on B cell function
2001
[34]
논문
Epstein–Barr virus (EBV)-associated epithelial and non-epithelial lesions of the oral cavity
2017
[35]
논문
Epstein–Barr virus and virus human protein interaction maps
2007-05
[36]
논문
Expression and processing of a small nucleolar RNA from the Epstein-Barr virus genome
2009-08
[37]
논문
Microbiome and malignancy
2011-10
[38]
논문
Constitutive interferon-inducible protein 16-inflammasome activation during Epstein–Barr virus latency I, II, and III in B and epithelial cells
2013-08
[39]
서적
Epstein–Barr Virus: Latency and Transformation
Caister Academic Press
[40]
논문
Latency and lytic replication in Epstein–Barr virus-associated oncogenesis
https://www.zora.uzh[...]
2023-04-19
[41]
논문
Stable replication of plasmids derived from Epstein-Barr virus in various mammalian cells
[42]
논문
Epstein–Barr virus: A master epigenetic manipulator
2017-10
[43]
논문
Eradication of Epstein–Barr virus by allogeneic bone marrow transplantation: implications for sites of viral latency
1988-11
[44]
논문
Epstein Barr virus (EBV) encoded small RNAs: Targets for detection by ''in situ'' hybridisation with oligonucleotide probes
1992-07
[45]
논문
Genome-wide analyses of Epstein–Barr virus reveal conserved RNA structures and a novel stable intronic sequence RNA
2013-08
[46]
논문
The role of microRNAs in Epstein–Barr virus latency and lytic reactivation
2011-12
[47]
논문
A bridge crosses the active-site canyon of the Epstein–Barr virus nuclease with DNase and RNase activities
2009-08
[48]
논문
Assessment of rapid ELISA test for detection of Epstein–Barr virus infection.
1990-08
[49]
논문
EBV Viral Loads in Diagnosis, Monitoring, and Response Assessment
2019-02-12
[50]
논문
Benign lymphadenopathies
2013-01
[51]
논문
Chronic fatigue syndrome: A manifestation of Epstein–Barr virus infection?
1988
[52]
논문
Long COVID or post-acute sequelae of COVID-19 (PASC): An overview of biological factors that may contribute to persistent symptoms
2021-06-23
[53]
논문
The role of EBV in the pathogenesis of Burkitt's Lymphoma: an Italian hospital based survey
[54]
논문
Epstein–Barr virus and hemophagocytic lymphohistiocytosis
[55]
논문
Epstein–Barr virus-associated Hodgkin's lymphoma
2004-05
[56]
간행물
Developing a vaccine for the Epstein–Barr virus could prevent up to 200,000 cancers globally say experts
https://www.cancerre[...]
2014-03-24
[57]
논문
Epigenetic dysregulation in Epstein–Barr virus-associated gastric carcinoma: Disease and treatments
2014-06
[58]
논문
Human papillomavirus and Epstein–Barr virus in nasopharyngeal carcinoma in a low-incidence population
2014-04
[59]
논문
Epstein–Barr virus genetic variants are associated with multiple sclerosis
2015-03
[60]
논문
Molecular signature of Epstein–Barr virus infection in MS brain lesions
2018-07
[61]
논문
Lymphomatoid granulomatosis: A practical review for pathologists dealing with this rare pulmonary lymphoproliferative process
2013-08
[62]
뉴스
Is a virus we all have causing multiple sclerosis?
https://www.bbc.com/[...]
2022-04-29
[63]
논문
Epstein–Barr virus and skin manifestations in childhood
2013-10
[64]
논문
Primary EBV infection and hypersensitivity to mosquito bites: a case report
2016-12
[65]
논문
The presence of Epstein–Barr virus significantly impacts the transcriptional profile in immunodeficiency-associated Burkitt lymphoma
[66]
논문
Epstein–Barr virus-associated lymphomas
2017-10
[67]
논문
Primary effusion lymphoma: Current concepts and management
2017-07
[68]
논문
Monoclonal antibodies against Epstein–Barr virus cross-react with alpha-synuclein in human brain
2000-11
[69]
논문
Epstein–Barr virus is an agent of genomic instability
https://www.nature.c[...]
2023-04
[70]
논문
Intra-host genomic diversity and integration landscape of human tissue-resident DNA virome
[71]
뉴스
Cancer virus discovery helped by delayed flight
https://www.bbc.co.u[...]
2015-11-04
[72]
논문
Virus Particles in Cultured Lymphoblasts from Burkitt's Lymphoma
1964-03
[73]
서적
Epstein–Barr Virus
Caister Academic Press
2010-09-18
[74]
서적
Epstein–Barr Virus
https://books.google[...]
Horizon Scientific Press
2012-06-03
[75]
book review
''Epstein–Barr Virus''
2006-12-21
[76]
논문
Epstein-Barr virus (EBV) reactivation and therapeutic inhibitors
2019-10
[77]
논문
Epidemiologic aspects of Epstein–Barr virus (EBV)-associated diseases
[78]
서적
Desk Encyclopedia of Human and Medical Virology
Academic Press
[79]
논문
Contribution of viral recombinants to the study of the immune response against the Epstein–Barr virus
2008-12
[80]
논문
An atlas of the Epstein–Barr virus transcriptome and epigenome reveals host-virus regulatory interactions
2012-08
[81]
논문
Main Targets of Interest for the Development of a Prophylactic or Therapeutic Epstein–Barr Virus Vaccine
[82]
논문
Long-term administration of valacyclovir reduces the number of Epstein–Barr virus (EBV)-infected B cells but not the number of EBV DNA copies per B cell in healthy volunteers
2009-11
[83]
논문
Antiviral agents for infectious mononucleosis (glandular fever)
[84]
웹사이트
ICTV Taxonomy history: ''Human gammaherpesvirus 4''
https://ictv.global/[...]
2019-01-10
[85]
서적
ヘルペスウイルス感染症
中外医学社
1996
[86]
간행물
Reactivation of Epstein–Barr virus from latency
https://www.ncbi.nlm[...]
2004
[87]
간행물
Spectrum of Epstein–Barr virus-related diseases: a pictorial review
https://www.ncbi.nlm[...]
2009
[88]
간행물
Epstein-Barr Virus Infection of Mammary Epithelial Cells Promotes Malignant Transformation
https://www.ncbi.nlm[...]
2016
[89]
간행물
Autoimmune disease: A role for new anti-viral therapies?
https://www.ncbi.nlm[...]
2011
[90]
간행물
CD8+ T-Cell Deficiency, Epstein-Barr Virus Infection, Vitamin D Deficiency, and Steps to Autoimmunity: A Unifying Hypothesis
https://www.ncbi.nlm[...]
2012
[91]
간행물
Evasion of affinity-based selection in germinal centers by Epstein-Barr virus LMP2A
https://www.ncbi.nlm[...]
2015
[92]
뉴스
自己免疫疾患の引き金となるウイルス因子を同定
http://resou.osaka-u[...]
リソウ
2015-08-25
[93]
뉴스
【坂口至徳の科学の現場を歩く】 誰もが感染ヘルペス…EBウイルスの因子を解明 阪大、自己免疫疾患の治療に道
https://www.sankei.c[...]
産経WEST
2015-08-28
[94]
간행물
Epstein-Barr Virus Lytic Reactivation Activates B Cells Polyclonally and Induces Activation-Induced Cytidine Deaminase Expression: A Mechanism Underlying Autoimmunity and Its Contribution to Graves' Disease
https://www.ncbi.nlm[...]
2017
[95]
간행물
EBウイルスはバセドウ病の発症や憎悪の最終因子となる
http://www.kahyo.com[...]
科学評論社
2016-09
[96]
간행물
Epstein–Barr virus in autoimmune diseases
https://www.ncbi.nlm[...]
2008
[97]
뉴스
“人気声優の死 異常なしがEBV感染症…難病指定訴え”
https://mainichi.jp/[...]
毎日新聞
2016-02-03
[98]
웹사이트
松来未祐日記
https://blog.excite.[...]
2018-07-13
[99]
웹사이트
病名が分からない苦しみと闘った女性|ザ!世界仰天ニュース
https://www.ntv.co.j[...]
2018-07-13
[100]
간행물
Recombinant gp350 vaccine for infectious mononucleosis: a phase 2, randomized, double-blind, placebo-controlled trial to evaluate the safety, immunogenicity, and efficacy of an Epstein-Barr virus vaccine in healthy young adults
https://www.ncbi.nlm[...]
2007
[101]
간행물
特集 基礎から拓くDDS創薬フロンティア 抗体ベンチャー(イーベック)
https://doi.org/10.2[...]
2012
[102]
뉴스
Cancer virus discovery helped by delayed flight
https://www.bbc.com/[...]
BBC News, Health
2014
[103]
간행물
Virus particles in cultured lymphoblasts from Burkitt's lymphoma
1964
[104]
서적
1. The origins of EBV research: discovery and characterization of the virus
https://books.google[...]
Trowbridge: Cromwell Press
2005
[105]
서적
Epstein–Barr Virus
https://books.google[...]
Horizon Scientific Press
2005
[106]
논문
"Book Review: Epstein–Barr Virus"
New England Journal of Medicine
2006
[107]
논문
"Epidemiologic aspects of Epstein–Barr virus (EBV)-associated diseases"
https://www.ncbi.nlm[...]
Annals of the New York Academy of Sciences
1980
[108]
서적
Desk Encyclopedia of Human and Medical Virology
Academic Press
2009
[109]
논문
"Epstein-Barr virus infections in males with the X-linked lymphoproliferative syndrome"
https://www.ncbi.nlm[...]
Annals of Internal Medicine
1987
[110]
논문
"Oral manifestations in the era of HAART"
https://www.ncbi.nlm[...]
Journal of the National Medical Association
2003
[111]
논문
"Epstein-Barr virus in AIDS-related primary central nervous system lymphoma"
https://www.ncbi.nlm[...]
The Lancet
1991
[112]
논문
Detection of Epstein Barr virus in an hepatic leiomyomatous neoplasm in an adult human immunodeficiency virus 1-infected patient
https://www.ncbi.nlm[...]
1994
[113]
논문
Epstein Barr virus-associated smooth muscle tumour (EBV-SMT) of the urinary bladder
https://www.ncbi.nlm[...]
2017-11-22
[114]
논문
The association of Epstein-Barr virus with smooth-muscle tumors occurring after organ transplantation
https://www.ncbi.nlm[...]
1995-01-05
[115]
논문
Association of Epstein-Barr virus with leiomyosarcomas in young people with AIDS
https://www.ncbi.nlm[...]
1995-01-05
[116]
논문
Undifferentiated carcinoma of the salivary gland in Greenlandic Eskimos: demonstration of Epstein-Barr virus DNA by in situ nucleic acid hybridization
https://www.ncbi.nlm[...]
1991-08
[117]
논문
"Association of Epstein Barr virus infection (EBV) with breast cancer in rural Indian women"
https://www.ncbi.nlm[...]
ProS One
2009
[118]
논문
"Joint effects of Epstein-Barr virus and polymorphisms in interleukin-10 and interferon-γ on breast cancer risk"
https://www.ncbi.nlm[...]
The Journal of Infectious Disease
2012
[119]
논문
"Multiplex PCR/mass spectrometry screening of biological carcinogenic agents in human mammary tumors"
https://www.ncbi.nlm[...]
Journal of Clinical Virology
2014
[120]
논문
"Frequency and genome load of Epstein-Barr virus in 509 breast cancers from different geographical areas"
https://www.ncbi.nlm[...]
British Journal of Cancer
2001
[121]
논문
"Investigation of Epstein-Barr virus in breast carcinomas in Tunisia"
https://www.ncbi.nlm[...]
Pathology - Research and Practice
2011
[122]
논문
"Epstein–Barr virus in epithelial cell tumors: a breast cancer study"
https://www.ncbi.nlm[...]
Cancer Research
1995
[123]
논문
"Epstein–Barr virus as a marker of biological aggressiveness in breast cancer"
https://www.ncbi.nlm[...]
British Journal of Cancer
2011
[124]
논문
"Epstein–Barr virus infection and clinical outcome in breast cancer patients correlate with immune cell TNF-alpha/IFN-gamma response"
https://www.ncbi.nlm[...]
BMC Cancer
2014
[125]
논문
"Infection of autoreactive B lymphocytes with EBV, causing chronic autoimmune diseases"
https://www.ncbi.nlm[...]
Trends in Immunology
2003
[126]
논문
"The essential role of Epstein-Barr virus in the pathogenesis of multiple sclerosis"
https://www.ncbi.nlm[...]
The Neuroscientist
2011
[127]
논문
HLA-DR4 antigen and idiopathic dilated cardiomyopathy susceptibility: a meta-analysis involving 11,761 subjects
https://www.ncbi.nlm[...]
2011-02
[128]
논문
Human leukocyte antigen polymorphisms in Italian primary biliary cirrhosis: a multicenter study of 664 patients and 1992 healthy controls
https://www.ncbi.nlm[...]
2008-12
[129]
논문
Genetics of inflammatory bowel disease: the role of the HLA complex
https://www.ncbi.nlm[...]
2006-06-21
[130]
논문
"Autoantibodies from patients with systemic lupus erythematosus bind a shared sequence of SmD and Epstein-Barr virus-encoded nuclear antigen EBNA I"
https://www.ncbi.nlm[...]
European Journal of Immunology
1993
[131]
논문
"Potential role of the Epstein-Barr virus in systemic lupus erythematosus autoimmunity"
https://www.ncbi.nlm[...]
Clinical & Experimental Rheumatology
1998
[132]
논문
"Epstein-Barr virus and molecular mimicry in systemic lupus erythematosus"
https://www.ncbi.nlm[...]
Autoimmunity
2006
[133]
논문
"関節リウマチの原因としてのEpstein-Barr ウイルス"
https://doi.org/10.4[...]
日大医誌
2014
[134]
논문
"Aryl hydrocarbon receptor-mediated induction of EBV reactivation as a risk factor for Sjogren's syndrome"
https://www.ncbi.nlm[...]
Journal of immunology
2012
[135]
논문
"Epstein–Barr Virus Infection and Multiple Sclerosis: A Review"
https://www.ncbi.nlm[...]
Journal of Neuroimmune Pharmacology
2010
[136]
논문
"Absence of Epstein-Barr virus in the brain and CSF of patients with multiple sclerosis"
https://www.ncbi.nlm[...]
Neurology
2010
[137]
논문
"Epstein-Barr virus infection is not a characteristic feature of multiple sclerosis brain"
https://www.ncbi.nlm[...]
Brain
2009
[138]
논문
"Absence of Epstein-Barr virus RNA in multiple sclerosis as assessed by in situ hybridisation"
https://www.ncbi.nlm[...]
Journal of Neurology, Neurosurgery & Psychiatry
1994
[139]
논문
"Epstein-Barr virus in the multiple sclerosis brain: a controversial issue--report on a focused workshop held in the Centre for Brain Research of the Medical University of Vienna, Austria"
https://www.ncbi.nlm[...]
Brain
2011
[140]
논문
"Epstein-Barr virus and multiple sclerosis: Updating Pender's hypothesis"
https://www.ncbi.nlm[...]
Multiple Sclerosis and Related Disorders
2017
[141]
논문
"Defective T-cell control of Epstein-Barr virus infection in multiple sclerosis"
https://www.ncbi.nlm[...]
Clinical & Translational Immunology
2017
[142]
논문
"The influence of Epstein-Barr virus reactivation in patients with Graves' disease"
https://www.ncbi.nlm[...]
Viral Immunology
2011
[143]
논문
"Reactivation of persistent Epstein-Barr virus (EBV) causes secretion of thyrotropin receptor antibodies (TRAbs) in EBV-infected B lymphocytes with TRAbs on their surface"
https://www.ncbi.nlm[...]
Autoimmunity
2015
[144]
논문
"Presence of Epstein-Barr virus-infected B lymphocytes with thyrotropin receptor antibodies on their surface in Graves' disease patients and in healthy individuals"
https://www.ncbi.nlm[...]
Autoimmunity
2015
[145]
논문
"Production of thyrotropin receptor antibodies in acute phase of infectious mononucleosis due to Epstein-Barr virus primary infection: a case report of a child"
https://www.ncbi.nlm[...]
Springerplus
2015
[146]
논문
"Thyrotropin Receptor Antibody (TRAb)-IgM Levels Are Markedly Higher Than TRAb-IgG Levels in Graves' Disease Patients and Controls, and TRAb-IgM Production Is Related to Epstein-Barr Virus Reactivation"
https://www.ncbi.nlm[...]
Viral Immunology
2016
[147]
논문
Elevated serum immunoglobulin G4 levels in patients with Graves' disease and their clinical implications
https://www.ncbi.nlm[...]
2014-04-01
[148]
논문
Immunoglobulin G4: an odd antibody
https://www.ncbi.nlm[...]
2009-04-01
[149]
논문
What is IgG4? A review of the biology of a unique immunoglobulin subtype
https://www.ncbi.nlm[...]
2011-01-01
[150]
논문
Epstein-Barr virus specific diagnostic tests in infectious mononucleosis
https://www.ncbi.nlm[...]
1974-09-01
[151]
논문
Epstein-Barr virus-encoded small RNA induces IL-10 through RIG-I-mediated IRF-3 signaling
https://www.ncbi.nlm[...]
2008-07-10
[152]
서적
Janeway's immunobiology
https://www.worldcat[...]
[153]
논문
Lifetime of plasma cells in the bone marrow
https://www.ncbi.nlm[...]
1997-07-10
[154]
서적
The immune system
https://www.worldcat[...]
Garland Science
2009
[155]
논문
The binding affinity of human IgG for its high affinity Fc receptor is determined by multiple amino acids in the CH2 domain and is modulated by the hinge region
https://www.ncbi.nlm[...]
1991-06-01
[156]
논문
Anti-inflammatory activity of human IgG4 antibodies by dynamic Fab arm exchange
https://www.ncbi.nlm[...]
2007-09-14
[157]
논문
Structural features of human immunoglobulin G that determine isotype-specific differences in complement activation
https://www.ncbi.nlm[...]
1993-08-01
[158]
논문
Epstein-Barr Virus Lytic Reactivation Induces IgG4 Production by Host B Lymphocytes in Graves' Disease Patients and Controls: A Subset of Graves' Disease Is an IgG4-Related Disease-Like Condition
https://www.ncbi.nlm[...]
2018-09-17
[159]
논문
Epstein-Barr virus infection of the colon with inflammatory bowel disease
https://www.ncbi.nlm[...]
1999-06-01
[160]
논문
Human cytomegalovirus and Epstein-Barr virus infection in inflammatory bowel disease: need for mucosal viral load measurement
https://www.ncbi.nlm[...]
2015-02-14
[161]
논문
Differential cellular localization of Epstein-Barr virus and human cytomegalovirus in the colonic mucosa of patients with active or quiescent inflammatory bowel disease
https://www.ncbi.nlm[...]
2016-02-01
[162]
논문
Looking into Enteric Virome in Patients with IBD: Defining Guilty or Innocence?
https://www.ncbi.nlm[...]
2017-08-01
[163]
논문
Refractory inflammatory bowel disease: is there a role for Epstein-Barr virus? A case controlled study using highly sensitive EBV encoded small RNA1 In Situ hybridization
https://www.ncbi.nlm[...]
2018-08-15
[164]
논문
PCR detection of human herpesviruses in colonic mucosa of individuals with inflammatory bowel disease: Comparison with individuals with immunocompetency and HIV infection
https://www.ncbi.nlm[...]
2017-01-01
[165]
논문
Combined Infection of Human Herpes Viruses: A Risk Factor for Subsequent Colectomy in Ulcerative Colitis
https://www.ncbi.nlm[...]
2018-05-18
[166]
논문
A novel interleukin-12 p40-related protein induced by latent Epstein-Barr virus infection in B lymphocytes
https://www.ncbi.nlm[...]
1996-02-01
[167]
논문
The expression of IL-12 p40 and its homologue, Epstein-Barr virus-induced gene 3, in inflammatory bowel disease
https://www.ncbi.nlm[...]
2001-08-01
[168]
논문
Epstein-Barr virus (EBV) infection and expression of the interleukin-12 family member EBV-induced gene 3 (EBI3) in chronic inflammatory bowel disease
https://www.ncbi.nlm[...]
2004-07-01
[169]
논문
Interleukin 27 limits autoimmune encephalomyelitis by suppressing the development of interleukin 17-producing T cells
https://www.ncbi.nlm[...]
2006-09-01
[170]
논문
Expression of Epstein-Barr virus-induced gene 3 and other interleukin-12-related molecules by human intestinal epithelium
https://www.ncbi.nlm[...]
2004-07-01
[171]
논문
The inhibitory cytokine IL-35 contributes to regulatory T-cell function
https://www.ncbi.nlm[...]
2007-11-22
[172]
논문
IL-35 is a novel cytokine with therapeutic effects against collagen-induced arthritis through the expansion of regulatory T cells and suppression of Th17 cells
https://www.ncbi.nlm[...]
2007-11-01
[173]
논문
Interleukin-35 mediates mucosal immune responses that protect against T-cell-dependent colitis
https://www.ncbi.nlm[...]
2011-11-01
[174]
논문
Risk of malignant lymphoma in patients with inflammatory bowel diseases: a Dutch nationwide study
https://www.ncbi.nlm[...]
2011-09-01
[175]
논문
Excess primary intestinal lymphoproliferative disorders in patients with inflammatory bowel disease
https://www.ncbi.nlm[...]
2012-11-01
[176]
논문
Risk factors for lymphoma in patients with inflammatory bowel disease: a case-control study
https://www.ncbi.nlm[...]
2013-06
[177]
논문
Epstein-Barr virus in inflammatory bowel disease: the spectrum of intestinal lymphoproliferative disorders
https://www.ncbi.nlm[...]
2015-05
[178]
논문
XIAP variants in male Crohn's disease
https://www.ncbi.nlm[...]
2015-01
[179]
간행물
【話題の感染症】 慢性活動性EBウイルス感染症
https://www.eiken.co[...]
2010
[180]
논문
CD40 ligand is a critical effector of Epstein-Barr virus in host cell survival and transformation
https://www.ncbi.nlm[...]
2003
[181]
논문
Coexpression of CD40 and CD40 ligand in Epstein-Barr virus-infected T and NK cells and their role in cell survival
https://www.ncbi.nlm[...]
2005
[182]
논문
“Monoclonal antibodies against Epstein-Barr virus cross-react with alpha-synuclein in human brain”
https://www.ncbi.nlm[...]
2000
[183]
논문
Alice in Wonderland syndrome as an initial manifestation of Epstein-Barr virus infection
https://www.ncbi.nlm[...]
1992-05
[184]
논문
"Alice in Wonderland" syndrome as a presenting symptom of infectious mononucleosis in children: a description of three affected young people
https://www.ncbi.nlm[...]
1977-02
[185]
논문
核磁気共鳴画像で異常を認めた EB ウイルス脳症による不思議の国のアリス症候群の 1 例
2002
[186]
논문
Epstein-Barr virus-associated acute renal failure: diagnosis, treatment, and follow-up
https://www.ncbi.nlm[...]
2003-07
[187]
논문
A cohort study among university students: identification of risk factors for Epstein-Barr virus seroconversion and infectious mononucleosis
https://www.ncbi.nlm[...]
2006-08-01
[188]
논문
Phase I trial of a CD8+ T-cell peptide epitope-based vaccine for infectious mononucleosis
https://www.ncbi.nlm[...]
2008-02
[189]
논문
Phase I/II studies to evaluate safety and immunogenicity of a recombinant gp350 Epstein-Barr virus vaccine in healthy adults
https://www.ncbi.nlm[...]
2007-06-11
[190]
논문
Dual stimulation of Epstein-Barr Virus (EBV)-specific CD4+- and CD8+-T-cell responses by a chimeric antigen construct: potential therapeutic vaccine for EBV-positive nasopharyngeal carcinoma
https://www.ncbi.nlm[...]
2004-01
[191]
논문
Epstein-Barr virus particles induce centrosome amplification and chromosomal instability
https://www.ncbi.nlm[...]
2017-02-10
[192]
논문
Structure-activity relationship between (E)-5-(2-bromovinyl)- and 5-vinyl-1-beta-D-arabinofuranosyluracil (BV-araU, V-araU) in inhibition of Epstein-Barr virus replication
https://www.ncbi.nlm[...]
1992-01
[193]
논문
Comparison of two bromovinyl nucleoside analogs, 1-beta-D-arabinofuranosyl-E-5-(2-bromovinyl)uracil and E-5-(2-bromovinyl)-2'-deoxyuridine, with acyclovir in inhibition of Epstein-Barr virus replication
https://www.ncbi.nlm[...]
1988-07
[194]
웹사이트
ソリブジン物語:この世から消えた有用な医薬品
https://kusuri-jouho[...]
2018-07-13
[195]
논문
Progress and Problems in Understanding and Managing Primary Epstein–Barr Virus Infections
https://www.ncbi.nlm[...]
2011
[196]
논문
Epstein Barr virus entry; kissing and conjugation
https://www.ncbi.nlm[...]
2014
[197]
논문
Epstein–Barr virus lacking glycoprotein gp42 can bind to B cells but is not able to infect
https://www.ncbi.nlm[...]
1998
[198]
논문
Human complement receptor type 1/CD35 is an Epstein–Barr Virus receptor
https://www.ncbi.nlm[...]
2013
[199]
논문
(-)-Epigallocatechin-3-gallate inhibition of Epstein-Barr virus spontaneous lytic infection involves ERK1/2 and PI3-K/Akt signaling in EBV-positive cells
https://www.ncbi.nlm[...]
2013
[200]
논문
Epstein–Barr virus and virus human protein interaction maps
https://www.ncbi.nlm[...]
2007
[201]
논문
Expression and processing of a small nucleolar RNA from the Epstein-Barr virus genome
https://www.ncbi.nlm[...]
2009
[202]
서적
Epstein–Barr Virus: Latency and Transformation
Caister Academic Press
2010
[203]
논문
EBウイルスの感染様式とがん
https://doi.org/10.2[...]
2014
[204]
논문
Stable replication of plasmids derived from Epstein–Barr virus in various mammalian cells
https://www.ncbi.nlm[...]
1985
[205]
논문
Eradication of Epstein–Barr virus by allogeneic bone marrow transplantation: implications for sites of viral latency
https://www.ncbi.nlm[...]
1988
[206]
문서
Epstein-Barr virus(EBV)
http://www.ft-patho.[...]
[207]
논문
Mimicry of CD40 signals by Epstein-Barr virus LMP1 in B lymphocyte responses
https://www.ncbi.nlm[...]
1999
[208]
논문
Epstein-Barr Virus Lytic Reactivation Activates B Cells Polyclonally and Induces Activation-Induced Cytidine Deaminase Expression: A Mechanism Underlying Autoimmunity and Its Contribution to Graves' Disease
https://www.ncbi.nlm[...]
2017
[209]
논문
Epstein-Barr virus sustains Burkitt's lymphomas and Hodgkin's disease
https://www.ncbi.nlm[...]
2004
[210]
서적
Janeway's men'eki seibutsugaku
https://www.worldcat[...]
Nankōdō
2010
[211]
논문
EB virus-induced B lymphocyte cell lines producing specific antibody
https://www.ncbi.nlm[...]
1977-09-29
[212]
논문
In vitro production of human antibodies specifically reactive with human gastric cancer cells of established lines and autologous tissues
https://www.ncbi.nlm[...]
1987-11
[213]
웹인용
ICTV Taxonomy history: ''Human gammaherpesvirus 4''
https://talk.ictvonl[...]
2019-01-10
[214]
저널
Epstein-Barr virus (EBV)-associated lymphoid proliferations, a 2018 update
2018-09
[215]
저널
Spectrum of Epstein-Barr virus-related diseases: a pictorial review
2009-01
[216]
저널
Oral manifestations in the era of HAART
2003-02
[217]
저널
Alice in Wonderland Syndrome: A Clinical and Pathophysiological Review
2016
[218]
저널
Post-infectious acute cerebellar ataxia in children
https://archive.org/[...]
2003-09
[219]
저널
Autoimmune disease: A role for new anti-viral therapies?
2011-12
[220]
저널
CD8+ T-Cell Deficiency, Epstein-Barr Virus Infection, Vitamin D Deficiency, and Steps to Autoimmunity: A Unifying Hypothesis
[221]
저널
Epstein-barr virus infection and multiple sclerosis: a review
2010-09
[222]
저널
Molecular signature of Epstein-Barr virus infection in MS brain lesions
2018-07
[223]
저널
Epstein-Barr virus is present in the brain of most cases of multiple sclerosis and may engage more than just B cells
2018
[224]
저널
Epstein-Barr virus in autoimmune diseases
2008-10
[225]
웹인용
Developing a vaccine for the Epstein–Barr Virus could prevent up to 200,000 cancers globally say experts
http://www.cancerres[...]
2014-03-24
[226]
저널
Global burden of deaths from Epstein-Barr virus attributable malignancies 1990-2010
2014
[227]
저널
Reactivation of Epstein-Barr virus from latency
2004-11
[228]
웹사이트
About 90% of adults have antibodies that show that they have a current or past EBV infection.
https://www.cdc.gov/[...]
National Center for Infectious Diseases
2016-08-08
[229]
웹인용
Epstein–Barr Virus and Infectious Mononucleosis
https://www.cdc.gov/[...]
CDC
2011-12-29
[230]
저널
Is EBV persistence in vivo a model for B cell homeostasis?
1996-08
[231]
웹인용
About Epstein–Barr Virus (EBV)
https://www.cdc.gov/[...]
Centers for Disease Control and Prevention
2016-09-14
[232]
저널
Progress and problems in understanding and managing primary Epstein-Barr virus infections
2011-01
[233]
저널
Epstein Barr virus entry; kissing and conjugation
2014-02
[234]
저널
Epstein-Barr virus lacking glycoprotein gp42 can bind to B cells but is not able to infect
https://archive.org/[...]
1998-01
본 사이트는 AI가 위키백과와 뉴스 기사,정부 간행물,학술 논문등을 바탕으로 정보를 가공하여 제공하는 백과사전형 서비스입니다.
모든 문서는 AI에 의해 자동 생성되며, CC BY-SA 4.0 라이선스에 따라 이용할 수 있습니다.
하지만, 위키백과나 뉴스 기사 자체에 오류, 부정확한 정보, 또는 가짜 뉴스가 포함될 수 있으며, AI는 이러한 내용을 완벽하게 걸러내지 못할 수 있습니다.
따라서 제공되는 정보에 일부 오류나 편향이 있을 수 있으므로, 중요한 정보는 반드시 다른 출처를 통해 교차 검증하시기 바랍니다.
문의하기 : help@durumis.com